
Gastrula - what is it? Gastrulation (formation of the gastrula) and its methods The inner layer of the gastrula lining its cavity is called.
EMBRYOLOGY
Lesson No. 4
TOPIC: “Embryogenesis. Crushing, gastrulation"
CONTROL QUESTIONS
Definition of the concept of “crushing”.
The nature of zygote fragmentation in humans.
Dark and light blastomeres, their morphological characteristics, development potential.
Morula and blastocyst, their morphological characteristics.
Implantation of the embryo into the uterine mucosa. Stages of implantation.
Histiotrophic and hematotrophic types of embryonic nutrition.
Methods of gastrulation in humans.
Stages of gastrulation (delamination and immigration), their timing and mechanisms.
Factors influencing the mechanisms of gastrulation.
OBJECTIVE OF THE LESSON:
Understand the features and nature of zygote fragmentation in humans. Learn to distinguish between dark and light blastomeres. Understand the difference between a morula and a blastocyst. Know the stages of implantation and types of embryo nutrition depending on the stage of implantation. Study the methods of gastrulation in humans, know the stages and mechanisms of gastrulation.
The student must know:
the nature of zygote fragmentation in humans;
morphological characteristics of dark and light blastomeres, development potential;
morphological characteristics of the morula and blastocyst;
stages of embryo implantation into the uterine mucosa;
methods of gastrulation in humans, their timing and mechanisms.
The student must be able to:
on diagrams and tables be able to distinguish between dark and light blastomeres; differentiate morula and blastocyst;
distinguish the stages of embryo implantation into the uterine mucosa;
distinguish between methods of gastrulation in humans;
make sketches and draw up a written protocol.
SPLITTING UP
Splitting up is a series of mitotic divisions of the zygote to produce many daughter cells (blastomeres) smaller size. Mitotic divisions of the zygote and subsequently blastomeres occur with an increase in the number of cells, but without an increase in their mass, and are therefore called cleavage.
Types of crushing:
complete, or holoblastic, (lancelet, amphibians, mammals): the zygote is completely divided into blastomeres;
partial, or meroblastic (fish, reptiles, birds): only part of the zygote is crushed;
uniform: the resulting blastomeres are the same or similar in size;
uneven: blastomeres differ in size;
synchronous: blastomeres divide simultaneously;
asynchronous.
Based on the nature of the spatial arrangement of blastomeres, the following are distinguished: types of complete crushing:
radial: the resulting blastomeres are located above each other, creating a figure with radial symmetry (lancelet, amphibians);
spiral: overlying blastomeres are displaced relative to the underlying ones at an angle of 45°, thus arranged in a spiral (molluscs, nemerteans, annelids and some planarians);
bilateral (bilaterally symmetrical): in the early stages, the location of blastomeres occurs according to the law of bilateral symmetry, so that the blastomeres of the right side of the embryo correspond to exactly the same blastomeres of the left side (ascidians, nematodes and some other invertebrates)
chaotic (disordered): already after the third division there is no strict pattern in the arrangement of blastomeres; during further fragmentation, the resulting disorder intensifies (some jellyfish). External chaos, however, is regulated by internal mechanisms that have not yet been clarified, and leads to the formation of an animal of a specific species.
Partial crushing exists in two types:
discoidal: fragmentation, in which only part of the cytoplasm at the animal pole (fish, reptiles, birds) is divided into blastomeres;
superficial: the surface layer of the cytoplasm is crushed, and as it develops it is completely separated from the yolk (insects and most other arthropods).
During the process of crushing, the embryo moves along the fallopian tube and completes crushing in the uterine cavity on the 6th day of development. Cleavage ends with the formation of a blastula. This is a multicellular embryo, usually with a cavity inside. The blastula wall is called blastoderm, and the cavity - blastocoel(primary body cavity). In blastula it is also usually distinguished roof, bottom and delimiting them marginal zone. A blastula formed with complete uniform fragmentation is called uniform coeloblastula (lancelet), with complete uneven fragmentation - uneven coeloblastula, also called amphiblastula (amphibians) or blastocyst (mammals, humans), with partial discoid - discoblastula (reptiles, birds), with superficial - Steroblastula (coelenterates).
At the blastula stage, the polarity of the embryo is finally established, the degree of its integration increases, and exactly the order of arrangement of cells and the degree of their interaction arise that are necessary for directed cellular movements at the next stage of development - gastrulation.
At the late blastula stage in the lancelet and lower vertebrates, so-called presumptive areas are established, which contain material from certain organs and systems. The location and boundaries of presumptive areas, or presumptive primordia, were studied using the method of marking areas of the embryo with vital dyes (W. Vogt’s method, proposed in 1925). After applying a mark to the surface of the embryo, one can trace its movements and determine the fate of this area during gastrulation and further development.
The plan for the relative location of the presumptive rudiments of future organs is called the map of presumptive rudiments. The map of the rudiments, for example, in amphibians, accurately indicates the position of the areas from which the epidermis of the skin, organs of smell, hearing, the lens of the eye and others will develop.
Human embryo fragmentation complete, or holoblastic(cleavage furrows run through the entire embryo) , unevennew(as a result of crushing, blastomeres of unequal size are formed) and asynchronous(different blastomeres fragment at different rates, so the embryo at certain stages of fragmentation contains odd number cells). The first fragmentation (division) of the zygote is completed after 30 hours, resulting in the formation of two blastomeres covered with a fertilization membrane. The stage of two blastomeres is followed by the stage of three blastomeres.
From the very first divisions of the zygote, two types of blastomeres are formed - “dark” and “light”. “Light”, smaller, blastomeres are crushed faster and are located in one layer around the large “dark” ones, which end up in the middle of the embryo. From the surface “light” blastomeres, a trophoblast subsequently arises, connecting the embryo with the maternal organism and providing it with nutrition. The internal, “dark” blastomeres form the embryoblast, from which the embryonic body and extraembryonic organs (amnion, yolk sac, allantois) are formed.
Starting from the 3rd day, fragmentation proceeds faster, and on the 4th day the embryo consists of 7-12 blastomeres. After 50 - 60 hours, a dense cluster of cells forms - morula, and on the 3-4th day. formation begins blastocysts- a hollow bubble filled with liquid. The blastocyst moves through the fallopian tube to the uterus within 3 days and after 4 days enters the uterine cavity. The blastocyst is in the uterine cavity in a free form (free blastocyst) for 2 days (5th and 6th days). By this time, the blastocyst increases in size due to an increase in the number of blastomeres - embryoblast and trophoblast cells - to 100 and due to the increased absorption of uterine gland secretions by the trophoblast and the active production of fluid by trophoblast cells.
During the first 2 weeks of development, the trophoblast provides nutrition to the embryo due to the breakdown products of maternal tissues - histiotrophic type of nutrition. The embryoblast is located in the form of a nodule of germ cells (“germinal nodule”), which is attached internally to the trophoblast at one of the poles of the blastocyst.
IMPLANTATION
Implantation (ingrowth, rooting) - the introduction of an embryo into the mucous membrane of the uterus. There are two stages of implantation:
adhesion (sticking): the embryo attaches to the inner surface of the uterus;
invasion (immersion) - the introduction of the embryo into the tissue of the uterine mucosa.
On the 7th day. changes occur in the trophoblast and embryoblast associated with preparation for implantation:
the blastocyst retains the fertilization membrane;
in the trophoblast, the number of lysosomes with enzymes increases, ensuring the destruction (lysis) of the tissues of the uterine wall and thereby facilitating the introduction of the embryo into the thickness of its mucous membrane;
microvilli appearing in the trophoblast gradually destroy the fertilization membrane;
the germinal node flattens and turns into a germinal scutellum, in which preparation for the first stage of gastrulation begins.
Implantation lasts about 40 hours. Simultaneously with implantation begins gastrulation(formation of germ layers). This is the first critical period of development.
In the first stage the trophoblast attaches to the epithelium of the uterine mucosa, and two layers are formed in it - cytotrophoblast and symplastotrophoblast.
In the second stage symplastotrophoblast, producing proteolytic enzymes, destroys the uterine mucosa. The trophoblast villi that form in this case, penetrating into the uterus, successively destroy its epithelium, then the underlying connective tissue and vessel walls, and the trophoblast comes into direct contact with the blood of the maternal vessels. The embryo is nourished directly from the mother's blood (hematotrophic type of nutrition). From the mother's blood, the fetus receives not only all the nutrients, but also the oxygen necessary for breathing.
The hematotrophic type of nutrition, replacing the histiotrophic one, is accompanied by a transition to a qualitatively new stage of embryogenesis - second phasegastrulation and formation of extraembryonic organs.
GASTRULATION
The period of gastrulation is characterized by active movements of both individual cells of the embryo and cell masses, as a result of which three main layers of the body are formed in vertebrates, they are called germ layers:
ectoderm- outer germ layer;
mesoderm- middle germ layer;
endoderm- inner germ layer.
Germ layers in different animals are homologous entities And, i.e. during development they give identical structures: The ectoderm is always transformed into the outer covering of the body, and the lining of the midgut develops from the endoderm.
Other feature gastrulation period is that the resulting during cell division, unlike blastomeres, begin to grow and increase to the size of the mother, this happens active growth and an increase in the size of the embryo itself.
Gastrulation in humans occurs in two stages:
Stage I (delamination) occurs on the 7th day intrauterine development: from the material of the embryonic nodule (embryoblast), two leaves are formed: the outer leaf - epiblast(cells have the appearance of pseudostratified prismatic epithelium) and internal - hypoblast, facing the cavity of the blastocyst (small cubic cells with foamy cytoplasm).
Stage II (immigration) - on the 14-15th day of intrauterine development.
Methods of gastrulation, i.e., the mechanisms of formation of germ layers, differ in different animals and are largely determined by the structure of the blastula. There are four main methods of gastrulation:
intussusception (invagination);
delamination (stratification, splitting);
immigration (eviction);
epiboly (fouling).
As a result of gastrulation, an embryo appears - gastrula. The gastrula has a cavity - gastrocel(the cavity of the primary intestine), into which the opening leads - blastopore(primary mouth).
Depending on the further fate of the blastopore in development, all animals are divided into protostomes and deuterostomes. In protostomes, which include most invertebrates, a mouth opening is formed in place of the blastopore, and the opposite end becomes the posterior end of the body. In deuterostomes, which include chordates and some invertebrates, the blastopore develops into an anus or neurointestinal canal located at the posterior end of the body, and the oral opening erupts on the ventral side at the opposite end of the body. The blastopore is distinguished edges, or lips:dorsal, lateral and ventral.
An important result of gastrulation in chordates is the formation of the so-called axial complex of primordia within the germ layers. The axial complex of rudiments represents the rudiments of the nervous system (neural plate) and notochord (chordal plate) located along the axis of the body of the embryo, as well as the rudiments of mesoderm lying laterally in relation to the notochordal plate and associated with it. The closely adjacent rudiments of the notochord and mesoderm are often called chordomesodermis.
There are several gastrulation factors:
physical: these include the faster division of small cells compared to larger ones, which leads to the overgrowth of large cells with small ones, for example, in the blastula of amphibians and fish that have telolecithal eggs. In the blastula of these animals, there is a gradient in the distribution of blastomeres in size - their sizes decrease in the direction from the vegetative pole to the animal one. The metabolic gradient has the same direction - with a decrease in cell size, their metabolic activity and division rate increase. It is in areas of active cell division that the processes of cell movement begin. One of the reasons for cell movement is considered to be a change in surface tension with an increase in the number of cells. The cells are capable of performing active amoeboid movements;
chemical: the mechanism of inducing influence is associated with the release of special factors: proteins, nucleoproteins, steroids.
organizing centers: the direction of cell movement, and then their differentiation, are determined by induction - the influence of some areas or rudiments of the embryo on others. Such areas and rudiments are called organizing centers, or inductors. The theory of organizing centers, put forward by G. Spemann, establishes the presence of special areas in the embryo at different stages of its development that have an inducing effect on neighboring areas. Organizing centers, for example, include the anterior (dorsal) lip of the blastopore, which induces the formation of chordomesoderm, which, in turn, causes the differentiation of some ectoderm cells into the neural plate.
An important role in the regulation of intercellular interactions During development, specialized intercellular connections are made - gap contacts, appearing already during cleavage between blastomeres and, possibly, representing the first system of signal transmission between cells.
Types of crushing (check out)
A - radial crushing (lancelet);
B - bilateral crushing (ascaris);
IN - disordered crushing (flukes);
G - spiral crushing (clam) .
The numbers indicate the sequence of crushing stages.
Types of blastula (draw)

METHODS OF GASTRULATION (draw)

Cross section of a vertebrate embryo at the end of the second gastru phaselation (three germ layers and the axial complex of primordia) (draw)


Conceptus from fertilization to implantation.
Ovulation occurs on the 14th day of the menstrual cycle, fertilization occurs within 1 day. Three days from the moment of fertilization, the conceptus moves along the fallopian tube, on the 4th day it enters the uterine cavity and after 5.5–6 days is implanted into the endometrium. As it moves through the fallopian tube, fragmentation occurs, and a blastocyst appears in the uterine cavity, ready for implantation. 1 - oocyte immediately after ovulation, 2 - fertilization 12–24 hours after ovulation, 3 - stage of appearance of male and female pronuclei, 4 - beginning of fragmentation, 5 - two-cell stage (30 hours), 6 - morula, consisting of 12–16 blastomeres (3 days), 7 - advanced morula in the uterine cavity (4 days), 8 - early blastocyst (4.5–5 days), 9 - blastocyst implantation (5.5–6 days).

Compactization and morula in the early stages of development.
As a result of compaction, the size of the intercellular spaces between blastomeres decreases, they come closer together, and a morula is formed. The morula is divided into an internal part (cells connected by gap junctions) and an external part (cells connected by tight junctions). The embryoblast develops from the cells of the inner part of the morula, and the trophoblast is formed from the cells of the outer part. The inner cell mass and trophoblast are the main structural elements of the blastocyst.
HUMAN BLASTOCYST (draw)

Scheme of human blastocyst implantation on day 7.5 of development
(familiarize)

1 – epiblast;
2 – hypoblast;
3 – symplastotrophoblast;
4 – cytotrophoblast;
5 – amniotic sac;
6 – epithelium of the uterine mucosa;
7 – basal layer of the uterine mucosa;
8 – blood vessel;
9 - epithelium of the uterine gland.
The structure of a 2-week human embryo.
Scheme of the second stage of gastrulation (check out)

A - cross section of the embryo;
b - germinal disc (view from the amniotic sac)
chorionic epithelium;
chorionic mesenchyme;
lacunae filled with maternal blood;
secondary villus base;
amniotic leg;
amniotic sac;
yolk vesicle;
germinal shield during gastrulation;
primitive streak;
rudiment of intestinal endoderm;
vitelline epithelium;
epithelium of the amniotic membrane;
primary nodule;
prechordal process;
extraembryonic mesoderm;
extraembryonic ectoderm;
extraembryonic endoderm;
germinal ectoderm;
germinal endoderm.
Which is called gastrula, and the process of its formation is gastrulation.
The blastula, as a single-layer embryo, has not yet been differentiated into germ layers, or layers of cells. The embryo only acquires the characteristics of a multicellular animal when its body is divided into outer and inner germ layers - ecto- And endoderm. The ectoderm forms the primary covering of the body. The endoderm gives rise to the primary gut.
The concept of the germ layer was introduced by the famous natural scientist Karl Baer, who discovered the germ layers in the chicken embryo. He showed that in all vertebrates the formation of certain organs can be associated with three germ layers. The ectodermis forms the epidermis and its derivatives, such as hair, feathers, and nervous system and sensory epithelium. From the endoderm arise the intestines and associated organs, such as the liver and lungs. Third germ layer - mesoderm, forms the muscles, skeleton, excretory system and part of the gonads. Subsequently, it was proven that the theory of germ layers is quite applicable to the development of invertebrates, thus being universal. Of course, the germ layers are in fact not strictly specialized, since the boundaries between them can be violated due to the wide potential capabilities of the cells during individual development. At the same time, the main position of the theory of germ layers, according to which the basic structure of multicellular animals is consistent with two or three poorly differentiated rudiments, indicating the phylogenetic community of these animals, is completely justified.
So, the embryo acquires a metacellular level of development when its body is divided into ecto- and endoderm. This separation is achieved during the process of gastrulation.
The two-layer embryo at the vegetative pole forms the primary mouth, or blastopore, leading into the cavity of the primary intestine. Depending on the position of the primary mouth, two main groups are distinguished among bilaterally symmetrical animals: primary- And deuterostomes. In protostomes, the blastopore turns into the oral opening of the animal, while the anal opening arises from the secondarily folded ectoderm, connecting to the posterior region of the endodermal gut (Fig. 30, a). In deuterostomes, the primary mouth is transformed into the anus, and in the head region, the oral opening is re-formed in the form of an ectodermal invagination (Fig. 30, b).
Thus, the main processes occurring at this stage of embryogenesis are significant movements of cells relative to each other ( morphogenetic movements). As a result, an embryo appears with a complex anatomical structure.
Gastrulation is followed by a period during which both cell division and morphogenetic movements continue. At this time, the processes of cell differentiation and organogenesis are important. They vary greatly among representatives of different types of animals.
Methods of gastrulation (formation of a two-layer embryo - gastrula)
There are several ways to form a two-layer embryo - gastrula.
Immigration
The simplest way is the immigration (creeping) of some cells from the surface layer into the cavity of the blastula, their reproduction there and filling the entire blastocoel with a randomly located mass. The outer layer of cells is ectoderm, and the inner layer is endoderm (Fig. 29). In many lower multicellular organisms, two main structures are formed due to the internal layer: the epithelium of the midgut (the endoderm itself) and the surrounding tissues that make up the third germ layer, or mesoderm. These two layers (ento- and mesoderm), according to the proposal of I. I. Mechnikov, are called phagocytoblastoma, while the ectoderm is kinoblastoma. The functions of these layers are different.
Intussusception
In less primitive animals, the gastrula is formed not by the creeping of cells into the blastocoel, but by the screwing in of the outer epithelium, after which the screwed-in part becomes the endoderm. This process is called intussusception.
Delamination
If, after crushing the egg, the result is not a hollow ball, but a morula, then the two-layer structure is achieved by delamination (splitting). The essence of delamination is that the outer cells turn into epithelium, while the inner cells remain endoderm.
Epiboly (fouling)
Another method of forming a two-layer embryo is called epiboly or fouling. Epiboly is observed in the case of eggs rich in yolk, when future endoderm cells find themselves inside due to their overgrowth with cells of the animal pole. Material from the site
Evolution of gastrula
Embryologists and evolutionists give great importance the process itself that transforms a single-celled fertilized egg (zygote) into a multicellular two-layer embryo. But the theories of I.I. Mechnikov multicellular arose from spherical colonies of protozoa. Individual individuals of such a colony, having captured food, went to digest it into the cavity of the colony, then returning back. Over time, the division of cells into nutritional and motor cells, equipped with flagella, occurred. The colony ceased to be a hollow ball, since there were always nutritional cells inside, forming a phagocytoblast. Mechnikov called this structure of multicellular organisms parenchyma. Parenchy mula is a hypothetical primordial multicellular animal.
On the other hand, the no less famous zoologist E. Haeckel, again based on the observed processes occurring in the developing egg, suggested that the primary two-layer animal arose through invagination in a certain place of the protozoan colony-ball. Haeckel called this hypothetical animal gastrea.
What comes first - immigration or intussusception - is difficult to decide. But you should keep in mind general rule: if in one organism some process occurs through the movement of individual cells, and in another - from the bending of epithelial layers, the first organism is more primitive in this regard than the second. The fact is that intussusception requires that the body already has regulatory mechanisms that ensure friendly, coordinated behavior of invaginating cells.
On this page there is material on the following topics:
Gastrulation(from Latin gaster - stomach) - a complex process of chemical and morphological changes, which is accompanied by reproduction, growth, directed movement and differentiation of cells, resulting in the formation of germ layers - the sources of the primordia of tissues and organs, and complexes of axial organs.
At this stage of development of organisms, a two-layer embryo is formed - gastrula. In this case, two germ layers are formed - ectoderm (external) and endoderm (interior). The gastrula corresponds in structure to modern coelenterates. At the late stage of gastrulation, the third germ layer is formed - mesoderm (average).
These leaves subsequently give rise to embryonic rudiments, from which tissues and organs are formed. There are four types of gastrulation (Fig. 8).
Immigration(invasion) is the most primitive, initial form of gastrulation. All other types of gastrulation are derived from it. In this case, the blastoderm cells move into the blastocoel, where they settle on the inner surface and form endoderm , and the outer cells form ectoderm . In this case, the gastric cavity is formed - gastrocel – cavity of the primary intestine (coelenterates).
Intussusception(invagination) - the blastoderm at the vegetative pole bends inside the blastocoel and reaches the cells of the animal pole. In this case, a gastrocoel is formed, which communicates with the external environment through an opening - blastopore – primary mouth.
With the development of the blastopore, animals are divided into two groups:
protostomes– the blastopore turns into a real mouth (worms, mollusks, arthropods);
deuterostomes– the primary mouth turns into an anus at the posterior end of the body, and at the anterior the oral opening reappears (brachiopods, echinoderms, chordates).
Epiboly(fouling) - at the animal pole of the blastula, cells divide faster and creep onto large cells of the vegetative pole. From the cells of the animal pole it is formed ectoderm , and from the cells of the vegetative pole – endoderm. This type of gastrulation is characteristic of animals in which the egg contains an increased amount of yolk (cyclostomes, amphibians).
Delamination(stratification) - blastoderm cells divide, daughter cells move into the blastocoel, forming endoderm , and the outer cells form ectoderm . In this case, the blastopore is not formed, so the gastrocoel does not communicate with the external environment. This type of gastrulation is characteristic of animals that have lost large reserves of yolk in their eggs (coelenterates, higher placentals).
Fig.7. Genetic control of mammalian development (according to B.V. Konyukhov, 1976)
In ontogenetic development, there are periods of greatest sensitivity to the damaging effects of various factors. These periods are called critical periods of development. P.G. Svetlov postulated two general critical periods in the development of placental mammals. The first of them coincides with the process of implantation of the embryo (in humans, the end of the 1st - beginning of the 2nd week of pregnancy), the second - with the formation of the placenta (in humans, from the 3rd to the 6th week). According to other sources, the second critical period also includes the 7th and 8th weeks, when neurulation occurs and local organogenesis begins.
Damaging action during implantation usually leads to its disruption, early death fetus and its abortion. According to some data, 50–70% of fertilized eggs (embryos) do not survive the implantation period.
Developmental defects are deviations in the structure of an organ or an entire organism, leading to functional disorders: developmental defects are persistent morphological changes in an organ or the organism as a whole, beyond the limits of normal variations and arising in utero as a result of disturbances in the development of the embryo or fetus, sometimes after the birth of a child due to disruption of further organ formation. These changes cause disruption of the corresponding functions. Developmental anomalies are understood only as those defects in which anatomical changes do not lead to significant dysfunction, for example, deformations of the auricles that do not disfigure the patient’s face and do not significantly affect the perception of sounds. Severe developmental defects, in which the appearance of a child is disfigured, are often called deformities.
One of the mechanisms of gastrulation is invagination (invagination of part of the blastula wall into the embryo)
1 - blastula,
2 - gastrula.
The most simply structured gastrula is found in coelenterates - it is an ellipsoid-shaped embryo, in which the ectoderm is represented by an outer single-cell layer, and the endoderm is an internal accumulation of cells. A cavity is formed in the inner layer of the embryo (endoderm) - the so-called. "primary gut", or gastrocoel. Later, at the anterior end of the embryo, the so-called “primary mouth”, or blastopore, is the opening through which the primary gut communicates with the external environment.
The gastrula of sea urchins is considered to be a typical gastrula. It is formed by “invagination” of part of the surface of a spherical blastula. As a result of invagination, part of the blastoderm (blastula skin) is pressed inward and forms the gastrocoel (primary intestine). Gastrocoel cells belong to the endoderm. Part of the blastoderm remains on the surface of the embryo and forms the ectoderm. Some of the cells “move out” into the space between the outer layer of the embryo and the primary gut; these cells form the mesoderm. Also, the so-called intestines are separated from the primary intestine into the embryo. coelomic sacs, which are also part of the mesoderm. The opening through which intussusception occurs is the primary mouth (blastopore).
The human embryo passes through the gastrula stage on days 8-9 of development. The human gastrula is a flattened discoidal formation (the so-called “germinal disc”), which is formed from the “inner cell mass” of the blastocyst. The upper (that is, facing the animal pole) layer of the germinal disc is classified as ectoderm, the middle layer is referred to as mesoderm, the lower (that is, facing the vegetative pole, the future yolk sac) layer of the disc is classified as endoderm. The homolog of the primary gut in humans is the so-called. “primary yolk sac" - a space limited at the animal pole by the ectoderm of the germinal disc, and on other sides by the so-called. hypoblastoma - extra-embryonic endoderm.
The gastrula can be formed by invagination (intussusception or embolism) or by epiboly (for example, in some invertebrates). With epiboly, small ectodermal cells gradually become overgrown with large endodermal cells, and a cavity does not form immediately, but appears later.
In most animals, the embryo at the gastrula stage is not free-living, but is located in the egg membranes or in the uterus. But there are animals with free-swimming gastrula (for example, the free-swimming larva of Coelenterates - planula (parenchymula) - is a gastrula).
Evolutionary origin of gastrula
The presence of the gastrula stage in all multicellular animals serves as one of the proofs of the unity of their origin. According to the biogenetic Haeckel-Müller law, this circumstance indicates a common ancestor that existed in all multicellular animals, whose structure resembled the gastrula of modern animals. There are several hypotheses regarding the evolutionary origin of this hypothetical gastrula-like metazoan ancestor.
Ernst Haeckel in 1872 put forward the so-called. "gastrea theory". According to this hypothesis, the ancestors of all multicellular organisms were spherical multicellular colonies of flagellates (in resemblance to the blastula, Haeckel called this ancestral organism “blastea”), which swam in the sea as part of plankton and fed on small organic particles suspended in water (for example, bacteria). During evolution, the blastea underwent invagination (invagination) and formed an organism consisting of two layers of cells (external and internal), the inner layer of cells formed a “gut” that opened outward with a “mouth” (by similarity with the gastrula, Haeckel called this ancestral organism “gastrea” "). The biological meaning of the transformation of blastea into gastraea according to E. Haeckel was the specialization of cells. All blastea cells were the same; with the help of beating flagella, the cells supported the blastea in the water column, and also scooped up food particles for ingestion. Specialization occurred in the gastrea: the cells of the outer layer, using the beating of flagella, supported the gastrea in the water column, the cells of the inner layer, using the beating of the flagella, created a fluid current that pulled particles into the primary gut. The presence of a cavity in the gastrea gave it an evolutionary advantage - the gastrea, unlike the blastea, had the ability to eat food larger than the cells of the gastrea itself, since now the cells of the inner layer could secrete digestive enzymes into the gastric cavity. According to the gastrea theory, the most ancient type of gastrulation is intussusception; other types of gastrulation are secondary and appeared later in evolution. Thus, the most primitive form of gastrula, the planula, is a secondarily simplified embryonic form of coelenterates.
Ilya Ilyich Mechnikov in 1876-1886 formulated the so-called "phagocytella theory". According to this hypothesis, the evolution of the blastea did not proceed through invagination, but through the migration of cells of the outer layer into the spherical blastea. Mechnikov justified such eviction (“immigration”) as follows: blastea cells after capturing food particles (
The blastula, also called the germinal sac, is the final result of the cleavage process of the egg. The next stage, occupying an intermediate position between cleavage and organogenesis, in embryogenesis is gastrulation. Its main meaning is the formation of three germ layers: endoderm, ectoderm and mesoderm. In other words, it is with gastrulation that embryonic differentiation and morphogenesis of the organism begins.
Definition of the term "gastrulation"
Back in 1901, gastrulation was described as the path through which mesodermal, endodermal and ectodermal cells enter the embryo. This definition implies the presence of special organ-forming spaces in the blastula. Having understood this rather simple description, it is easy to move on to a more complex one, modern meaning term. Gastrulation is a sequence of morphogenetic movements, the result of which is the movement of tissue rudiments to places intended for them in accordance with the “plan” of the organization of the organism. The process is complex, changes are accompanied by growth and reproduction, directed movement and differentiation of cells.
Considering gastrulation in a more general sense, we can define it as an intermediate stage belonging to a single dynamic process, during which the blastula areas are rearranged, which significantly facilitates the transition to the process of organogenesis.
Moving Cells
If you give general characteristics process under consideration, then we can say that gastrulation is embolism and epiboly. Both terms reflect the morphogenetic movement of cells, which occurs at absolutely all stages of the ontogenetic development of the organism. However, they are most pronounced during gastrulation. Epiboly is the process of movement of cells along the surface of the embryo, and embolism is their movement inside it.
In embryology, the following main types of gastrulation or cell movement are distinguished: invagination, immigration, involution, delamination and epiboly. More details about them are provided later in the article.
Movement of cell layers
Not only individual (freely migrating) cells, but also entire cell layers can take part in the gastrulation process. The direction is determined by constant and distant interactions. The first forces were discovered by P. Weiss in the 20s of the last century and apparently also occur in embryogenesis, the second are rare and special, and occur with a low degree of probability during normal morphogenesis.
During gastrulation, cell fragmentation does not occur. As mentioned above, the movement of cell masses begins and, as a result, the formation of a two-layer embryo called a gastrula. The endoderm and ectoderm become clearly visible. In all multicellular organisms (the only exceptions are coelenterates), the third germ layer, called mesoderm, is formed in parallel with gastrulation or immediately after it. It is a collection of cells located between the ectoderm and endoderm. As a result, the embryo becomes three-layered.
The methods of gastrulation directly depend on the type of blastula.
Intussusception gastrula

The name of the method speaks for itself. Intussusception is the invagination of the single-layer wall of the blastula (balstoderm) into the blastocoel. The most primitive and most obvious example will be the one with a rubber ball. When you press it, part of the material is pressed inward. The invagination can be brought to the farthest wall or made insignificant. As a result, the blastula is transformed, and the gastrula is obtained in the form of a two-layer sac with an archenteron. Its inner wall is the primary endoderm, and its outer wall is the primary ectoderm. The resulting archenteron (primary intestine) communicates with the external environment through an opening called a blastopore. Its second name is the primary mouth. Its further development depends on the type of organism. In many animals, the blastopore eventually develops into a definitive mouth. In this regard, they are called protostomes (molluscs, worms, arthropods). In deuterostomes, the blastopore turns into a neurointestinal canal located in the posterior part of the embryo (in chordates), or into the anus.
Immigration gastrula
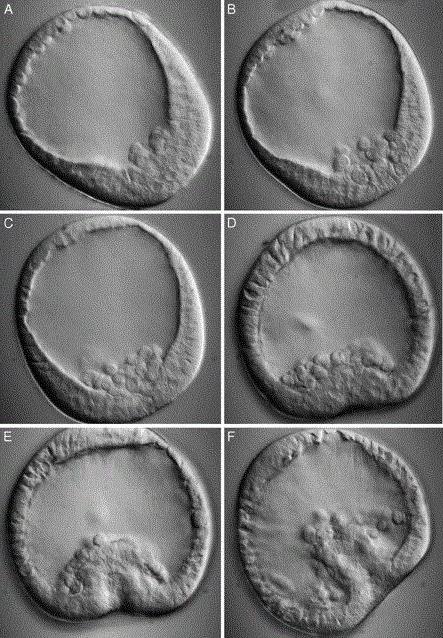
Immigration gastrulation is a method of formation of a two-layer embryo, most characteristic of coelenterates. The gastrula is formed by the active migration of part of the blastula cells into the blastocoel. Such immigration is unipolar. Cells move only from the vegetative pole. Later they form the endoderm, i.e. the inner layer. It is in this way that gastrulation occurs in the hydroid polyp, the jellyfish.
Blastodermal cells can penetrate into the blastocoel not in any one area, but throughout the entire surface of the embryo. This type of immigration is called multipolar, but it is quite rare.
In many coelenterates, which are characterized by the immigration method of gastrulation, a very active “eviction” of blastula cells occurs, and the resulting gastrula completely loses its blastocoel. In this case, the blastopore characteristic of the previous invagination method is absent.
Delamination gastrula

This rare type of gastrula was first described by I. I. Mechnikov, and it is characteristic of coelenterates. The processes accompanying gastrulation are very peculiar, but when considering a typical case they are perceived more simply. For example, the eggs of some scyphojellyfish have concentrically located and clearly distinguishable sections of cytoplasm: dense and granular (ectoplasm) and cellular (endoplasm). They are characterized by relatively synchronous and uniform division: 2, 4, 8, 16. Ultimately, the embryo contains 32 blastomeres. Further division occurs parallel to the surface of the embryo. An outer layer of blastomeres is formed, consisting of ectoplasm, and an inner layer - partly from ectoplasm and endoplasm. In other words, the process of formation of a multilayer embryo occurs by splitting one layer of cells into two. Then only the internal blastomeres are fragmented and again parallel to the surface of the embryo, which, as a result of such a peculiar gastrulation, takes on the shape of a ball. It consists of 64 flat cells forming the ectoderm and 32 more convex cells, which form the basis of the endoderm.
Epibolic gastrula

In animals with a pronounced telolecithal structure of eggs (displacement of the yolk towards the vegetative pole), gastrulation occurs according to the epibolic method. Macromeres are large blastomeres that divide very slowly and contain a large number of yolk. They lack the ability to move; therefore, more active micromeres located on the cell surface literally “crawl” onto them. With such gastrulation, the blastopore is absent and the archenteron is not formed. Only later, when the macromeres nevertheless decrease in size, does a cavity begin to form, the rudiment of the primary intestine.
Involution
Involutional gastrulation is a process that involves “tucking” the outer layer of cells inside the embryo. It increases in size and spreads along the inner surface. This method of gastrulation is characteristic of animals with mesolecithal eggs - amphibians (amphibians). The movement of the leading deep cells of the marginal region inhibits the development of the archenteron. It is in them that the driving force of involution lies.

Mixed method of gastrulation
As is known, embryogenesis is the earliest period of development of each individual organism: from conception to birth. Gastrulation is one of its stages, the second in chronology after cleavage. Its methods are so different that comparing them can be done with a great deal of convention. Each of them requires detailed study and analysis. However, there are still certain lines of intersection between them. Thus, the process of epiboly can be considered as a unique variant of intussusception, and delamination has features similar to immigration.
Note that in many animals gastrulation occurs in a combined way. In such cases, epiboly and invagination, as well as other morphogenetic processes, occur simultaneously. In particular, this is how gastrulation occurs in amphibians. In this regard, many authors highlight a mixed method.
Gastrula

Literally from Latin, the term “gastrula” is translated as “womb, stomach.” It denotes a specific embryo of multicellular organisms. Distinctive feature Gastrulae are the presence of two or three germ layers. The process of its formation is the gastrulation phase.
The simplest device is observed in animals. They are characterized by an ellipsoidal gastrula with a single-celled outer layer (ectoderm) and an internal accumulation of cells (endoderm), as well as a “primary gut”. The gastrula is considered typical sea urchin, which is formed by intussusception. In humans, gastrulation occurs on the 8th-9th day of development. The gastrula is a disc-shaped flattened formation formed from the inner cell mass.
As a rule, in most animals at the gastrula stage, the embryo cannot live freely and is located in the uterus or egg membranes. However, there are exceptions. Thus, the larvae of coelenterates, planulae, are a free-swimming gastrula.